
Разделы
- Главная страница
- Новости
- Краткий исторический экскурс
- Эра динозавров
- Гигантские растительноядные динозавры
- Устрашающие хищные динозавры
- Удивительные птиценогие динозавры
- Вооруженные рогами, шипами и панцирями
- Характерные признаки динозавров
- Загадка гибели динозавров
- Публикации
- Интересные ссылки
- Статьи
- Архив
Статистика
Д. Рауп, С. Стэнли. «Основы Палеонтологии»
Знаменитый палеонтолог XIX века Э. Коп, работавший преимущественно в области палеонтологии позвоночных, отметил общую тенденцию к увеличению размеров в ходе филогенеза в различных таксономических группах. Эту тенденцию нельзя считать ни ортогенетической, ни всеобщей, но тем не менее она широко распространена.
Мы уже говорили о тенденции к увеличению размеров, характерной для эволюции рогатых динозавров (фиг. 115). В общей картине эволюции лошадей наблюдается та же тенденция, как явствует из сравнения современного рода Equus и его четвертичных родичей с эоценовой ранней лошадью Hyracotherium, бывшей величиной примерно с фокстерьера. Увеличение размеров началось только после эоцена, а в некоторых эволюционных рядах вместо увеличения шло уменьшение. Несколько примеров явного увеличения размеров в филогенезе беспозвоночных рассмотрел Ньюэлл [133].
Причины увеличения размеров различны в разных эволюционных рядах. Некоторые животные благодаря увеличению размеров, возможно, становились сильнее и могли нападать на других животных или успешно защищаться. Теплопродукция у теплокровных животных пропорциональна объему их тела (т. е. кубу линейных размеров), а потеря тепла пропорциональна поверхности тела (т. е. квадрату линейных размеров). В результате крупное животное лучше сохраняет тепло в холодном климате, чем мелкое животное. Другое преимущество больших размеров состоит в том, что, хотя крупное животное бегает лишь ненамного быстрее мелкого, оно гораздо выносливее и может дольше бежать с большой скоростью. Мышцы крупного животного сокращаются медленнее, но зато размах каждого движения шире (например, шире шаг при беге). Поэтому примерно при одинаковой скорости передвижения крупное животное на том же расстоянии сделает меньше движений, чем мелкое, т. е. его выносливость опять-таки будет больше. Этот фактор мог иметь большое значение в эволюционном увеличении размеров лошадей, питавшихся травой.
Важно помнить, что с увеличением размеров связаны, и некоторые затруднения (это вытекает из принципа подобия). Например, увеличение размера травоядных должно было вызвать затруднения в добывании пищи, так как жевательная поверхность зубов увеличивалась пропорционально квадрату линейных размеров, а объем живых тканей, для поддержания которых Нужна пища, — пропорционально кубу линейных размеров. Симпсон интерпретирует сложные эволюционные изменения коренных зубов у лошади (фиг. 117) как адаптации, способствующие решению этой проблемы. В целом увеличение размеров — самоограничивающийся процесс. Человек тридцатиметрового роста не мог бы ходить и с трудом выдерживал бы собственный вес, поскольку вес увеличивается пропорционально кубу линейных размеров, а мышечная сила (которая пропорциональна площади сечения мышцы) — пропорционально их квадрату. Многие функции, связанные с различными поверхностями организма, при увеличении общих размеров также становятся менее эффективными. Например, потребность в пище растет вместе с объемом тканей, который пропорционален кубу линейных размеров, тогда как добывание пищи у таких организмов, как брахиоподы, мшанки, двустворчатые моллюски, зависит от снабженных ресничками Поверхностей, а их эффективность пропорциональна лишь квадрату размеров. Потребность в кислороде тоже возрастает пропорционально кубу линейных размеров, тогда как поглощение кислорода идет на двумерных поверхностях жабр или легких.
Можно взглянуть на правило Копа и по-другому: ведь здесь можно говорить не о тенденции к увеличению размеров в процессе эволюции, а об эволюции от малых размеров. Хорошо известно, что многие высшие таксоны развились из групп организмов, обладавших сравнительно малыми размерами. Во многом это объясняется тем, что предковые группы обычно бывают относительно неспециализированными, а мелкие особи гораздо чаще бывают неспециализированными, чем крупные. В большинстве случаев у видов, для которых характерны крупные размеры, возникло слишком много специальных адаптаций, и они вряд ли смогут дать начало большой новой группе с настолько важными отличиями, чтобы ее пришлось выделить в новый высший таксон. Например, очень маловероятно, чтобы слоны могли дать начало совершенно новому отряду четвероногих. Присущие слонам особые признаки, в том числе сильно утолщенные конечности, поддерживающие массивное тело, укороченная шея, несущая голову, и хобот, заменяющий длинную подвижную шею [162], значительно ограничивают их эволюционный потенциал. Новые группы обычно возникают не от крупных специализированных предков, а от мелких неспециализированных, и, может быть, именно поэтому правило Копа адекватно отражает действительность.
ПРИСПОСОБИТЕЛЬНОЕ ЗНАЧЕНИЕ МОРФОЛОГИЧЕСКИХ ТЕНДЕНЦИЙ
Приспособительное значение многих морфологических тенденций, отмечаемых в палеонтологической летописи, остается загадкой. В качестве примера можно привести изменение скульптуры раковины, обнаруженное Бринкманом в эволюционном ряду Kosmoceras (стр. 260). Беспозвоночные очень многочисленны в морских местообитаниях, и поэтому они чаще позвоночных оставляют в палеонтологической летописи почти непрерывную запись филетических изменений. Один из наиболее подробно исследованных ископаемых рядов—род морских ежей Micraster из меловых пластов «мела» Англии. Первым филетические тенденции Micraster выявил палеонтолог-любитель Роув [170]. Позже Кермак [100] подтвердил с помощью биометрических исследований основные наблюдения Роува, но отверг некоторые детали его работы. Совсем недавно Никольс [137, 138] проанализировал приспособительное значение найденных у Micraster морфологических тенденций. Совокупность работ этих палеонтологов — классический пример исследования эволюции.
Echinocardium — неправильный морской еж из группы морских ежей, сменивших исходную радиальную симметрию на билатеральную. Это изменение связано с однонаправленным передвижением животного по осадку или в его толще. Типичный современный роющий вид показан на фиг. 129. Он находится в незамкнутой норке и движется вперед, прорывая ход в осадке своими подвижными иглами, находящимися главным образом на брюшной поверхности. Связь с придонной водой поддерживается через вертикальную «воронку», проделанную особыми дыхательными амбулакральными ножками. Животное питается осадком, богатым органическим веществом, собирая этот осадок специальными амбулакральными ножками, окружающими рот. Некоторое количество пищи попадает в рот и с дорсальной поверхности, по передней борозде (фиг. 130). Иглы и амбулакральные ножки, окружающие анус, образуют задний санитарный канал, куда животное по мере своего продвижения выделяет экскременты.
На фиг. 130 показана морфология трех видов позднемелово го рода Micraster и трех современных неправильных морских ежей. Очень редко встречаются ископаемые морские ежи, у которых сохранились иглы, но о морфологии и расположении игл можно многое узнать, изучая шишковидные бугорки в местах прикрепления игл, так называемые туберкулы. Почти у всех видов сердцевидных морских ежей некоторые участки панциря несут ряды туберкул, к которым при жизни прикрепляются иглы, густо усаженные ресничками. Эти области называются фасциолами. Реснички внутренней фасциолы предназначены в основном для втягивания воды в дыхательную воронку. Реснички анальной фасциолы создают ток воды, относящий экскременты в сторону от тела и вниз, а реснички субанальной фасциолы помогают другим ресничкам тела гнать воду с продуктами выделения в санитарный канал.
![Фиг. 129. Echinocardium — типичный современный зарывающийся неправильный морской еж [137]](/images/osnpal/1-131.jpg "Фиг. 129. Echinocardium — типичный современный зарывающийся неправильный морской еж [137]")
Фиг. 129. Echinocardium — типичный современный зарывающийся неправильный морской еж [137]
На фиг. 131 показана предполагаемая глубина зарывания видов Micraster. Рассматриваемые виды соединены промежуточными формами и выделены условно. Предполагается, что исходный вид, М. leskei, зарывался неглубоко. Он дал начало ряду форм, среди которых были и глубоко, и неглубоко закапывавшиеся. Эту группу форм произвольно делят на два вида — М. corbovis и М. cortestudinarium. Только представители второго вида дожили до сенона и дали начало эволюционному ряду, составляющему вид М. coranguinum. Считается, что неглубоко закапывающийся вид М. (Isomicraster) senonensis появился в районе исследования в раннем сеионе, развился же он в географически изолированной области и поэтому долгое время не имел связи с основной популяцией. Он, видимо, не достиг той степени генетической изоляции, которая препятствовала бы скрещиванию с М. coranguinum (хотя палеонтолога рассматривают их как два отдельных вида); найдены промежуточные формы, свидетельствующие о скрещивании между этими двумя группами морских ежей.
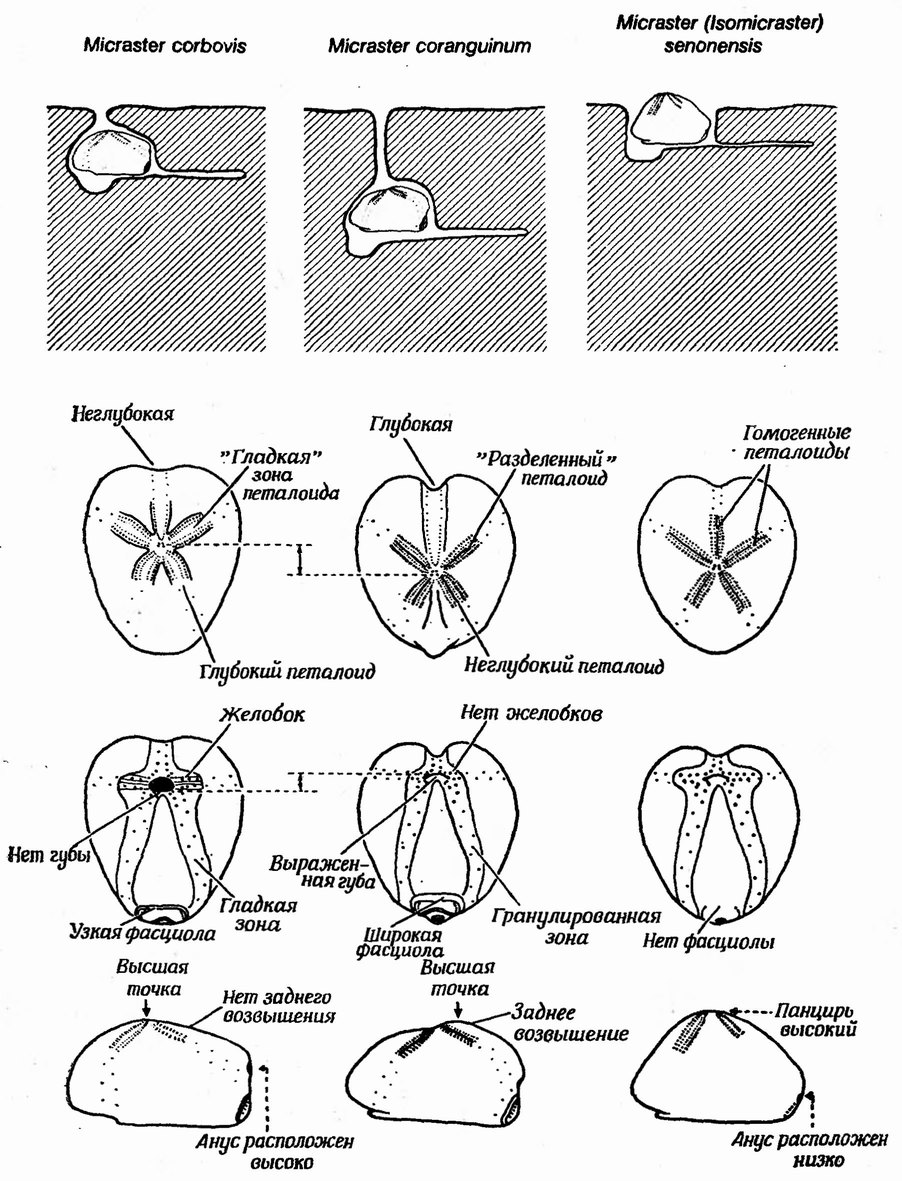
![Фиг. 130. Морфология и образ жизни современных рядом ископаемых неправильных морских ежей [138]](/images/osnpal/1-133.jpg "Фиг. 130. Морфология и образ жизни современных рядом ископаемых неправильных морских ежей [138]")
Фиг. 130. Морфология и образ жизни современных рядом ископаемых неправильных морских ежей [138]
По гомологии с рецентными формами и на основании адаптивной морфологии ископаемых Micraster Никольс сделал выводы об их образе жизни. Предполагаемая глубина зарывания трех видов Micraster и известная глубина зарывания трех современных ежей показаны на фиг. 130. Изучив некоторые приспособительные особенности, развившиеся в ряду М. cortestudinarium — М. coranguinum, Никольс выявил тенденцию к увеличению глубины зарывания. Ниже мы перечислим эти особенности (фиг. 130).
1. Расширение панциря, при котором самая широкая и высокая область сдвигается назад. Это изменение, очевидно, связано с приобретением обтекаемой формы и с отсутствием необходимости сбрасывать осадок с дорсальной поверхности (последнее важно для животных, которые лишь частично зарываются в донный осадок).
<< [1] ... [52] [53] [54] [55] [56] [57] [58] [59] [60] [61] [62] [63] ... [76] >>