
Разделы
- Главная страница
- Новости
- Краткий исторический экскурс
- Эра динозавров
- Гигантские растительноядные динозавры
- Устрашающие хищные динозавры
- Удивительные птиценогие динозавры
- Вооруженные рогами, шипами и панцирями
- Характерные признаки динозавров
- Загадка гибели динозавров
- Публикации
- Интересные ссылки
- Статьи
- Архив
Статистика
Д. Рауп, С. Стэнли. «Основы Палеонтологии»
Недостающее звено иногда удается выявить также благодаря открытию неизвестных прежде рецептных организмов. В качестве примера приведем глубоководного моллюска Neopilina. В 1952 г. участники датской экспедиции на «Галатее» подняли представителей этого необычного рода с океанского дна у Тихоокеанского побережья Центральной Америки, с глубины около 3500 м. Лемхе и Вингстранд [117] описали анатомию этого моллюска, которому присущи многие примитивные признаки. Наиболее необычен общий план строения: моллюск имеет ряды парных мышц ноги, органов, напоминающих почки, жабр (фиг. 147). У других групп моллюсков имеется только по одной паре этих органов, а если их по нескольку пар, то это вторичное (не примитивное) явление.
За два десятилетия, предшествовавших открытию Neopilina, некоторые палеонтологи, в том числе Венц и Найт, предполагали, что раннепалеозойские ископаемые, принадлежащие к классу Monoplacophora (фиг. 147), были блюдцеобразными предками брюхоногих, а может быть, и других высших моллюсков. Обнаруженная в 1952 г. Neopilina была отнесена к моноплакофорам, хотя ископаемые представители группы были известны только из отложений нижнего и среднего палеозоя. Neopilina— «живое ископаемое», представляющее группу, которая, видимо, на много миллионов лет нашла убежище на дне глубоководной части океана.
Эволюционное положение моноплакофор, анатомия которых известна только но Neopilina, все еще является спорным. Во-первых, неясно, можно ли считать полусегментированное строение тела примитивных моноплакофор доказательством того, что моллюски произошли от сегментированных кольчатых червей. Уже давно аннелиды и моллюски считаются родичами из-за сходства ранних стадий их онтогенеза. Во-вторых, возник вопрос, действительно ли моноплакофоры, вроде Neopilina, дали начало высшим классам моллюсков. На большинстве схем, реконструирующих облик «первого моллюска», имеется только по одной паре каждого органа. Если Neopilina находится на главной линии ранней эволюции моллюсков, то возникновению высших классов моллюсков должно было предшествовать исчезновение сегментации. Нет веских доказательств того, что группа, к которой относится Neopilina, дала начало какому-либо другому классу моллюсков. Несмотря на свое неясное филогенетическое положение, Neopilina — один из самых важных живых таксонов, открытых в последние десятилетия.
Наиболее плодотворным для выяснения филогенезов оказывается совместное рассмотрение палеонтологических и неонтологических данных. Использование только палеонтологических или только неонтологических данных может привести к ошибкам. Мы не будем приводить конкретных примеров, но несостоятельность большинства гипотез палеонтологов и неонтологов о связях между таксономическими группами обусловлена именно использованием односторонней информации.
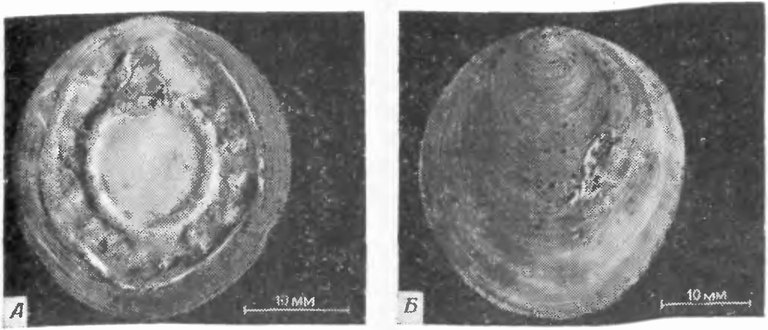
![Фиг. 147. Monoplacophora [117]. А—E — Современный вид Neopilina galatheae Lemche (А и В — брюшная сторона, Б и Г — спинная сторона, Д — боковая сторона, Е — вид спереди; 1 — ротовое отверстие, 2 — жабры, 3 — край ноги, 4 — край мантии, 5 — анус, 6 — край раковины). Ж. Силурийская форма Pilina unguis; видны отпечатки парных мышц, которыми прикреплялись тело и нога, на внутренней стенке раковины](/images/osnpal/1-152.jpg "Фиг. 147. Monoplacophora [117]. А—E — Современный вид Neopilina galatheae Lemche (А и В — брюшная сторона, Б и Г — спинная сторона, Д — боковая сторона, Е — вид спереди; 1 — ротовое отверстие, 2 — жабры, 3 — край ноги, 4 — край мантии, 5 — анус, 6 — край раковины). Ж. Силурийская форма Pilina unguis; видны отпечатки парных мышц, которыми прикреплялись тело и нога, на внутренней стенке раковины")
Фиг. 147. Monoplacophora [117]. А—E — Современный вид Neopilina galatheae Lemche (А и В — брюшная сторона, Б и Г — спинная сторона, Д — боковая сторона, Е — вид спереди; 1 — ротовое отверстие, 2 — жабры, 3 — край ноги, 4 — край мантии, 5 — анус, 6 — край раковины). Ж. Силурийская форма Pilina unguis; видны отпечатки парных мышц, которыми прикреплялись тело и нога, на внутренней стенке раковины
Эволюция экосистем
В истории жизни большую роль сыграли различные новшества, имевшие большое адаптивное значение. Многие из них имели лишь весьма небольшое значение для общей экологической истории Земли, тогда как другие были очень важны, поскольку прямо или косвенно приводили к новому образу жизни и к освоению новых местообитаний. Так появлялись новые экосистемы, и биосфера сильно изменилась с тех пор, как впервые появилась жизнь.
Видимо, первым важным событием в истории жизни было возникновение механизма самокопирования с помощью нуклеиновых кислот. К сожалению, по палеонтологической летописи трудно датировать это событие. С тех пор прошли сотни миллионов лет, на протяжении которых осадки в значительной мере изменялись.
Вторым важным событием в органической эволюции было возникновение клетки. Древнейшие остатки организмов, обладавших, по-видимому, клеточным строением, найдены в формации Онвервах.т в Южной Африке (3,2 млрд. лет). Эти остатки представляют собой мельчайшие палочковидные и сферические структуры, сохранившиеся в кремнистых сланцах и других мелкозернистых осадочных породах. Баргхурном и Шопфом [7J описаны лучше сохранившиеся палочковидные образования, напоминающие бактерий, и водорослевые сферы из формации Фиг-Три в Южной Африке (минимальный возраст 3,1 млрд. лет). Из древних докембрийских пород известны также строматолиты (однако широкое распространение они, видимо, получили только в среднем докембрии).
Наиболее полно изученные докембрийские отложения, содержащие остатки микроорганизмов — кремнистые сланцы формации Ганфлинт (Онтарио, Канада) (около 1,9 млрд. лет). Все микроорганизмы, описанные из этой формации, делятся на два основных типа: структуры, сходные с сине-зелеными водорослями (иногда эти структуры образуют строматолиты), и напоминающие бактерий структуры (фото XXXII). Сине-зеленые водоросли и бактерии отличаются от всех других современных клеточных организмов некоторыми примитивными чертами. Самая важная из них состоит в том, что генетический материал таких клеток не упакован в хромосомы в специальной клеточной структуре — ядре. Такие клетки, не имеющие хромосом и ядра, называются прокариотическими. Прокариотическая клетка может содержать большой набор генов, и одна мутация вряд ли может сильно изменить клетку, а поскольку прокариоты размножаются бесполым путем, мутации медленно распространяются в популяции. Этим может объясняться необычайно низкая скорость эволюции бактерий и сине-зеленых водорослей и сходство их докембрийских представителей с современными [184].
Хромосомы и оформленное ядро характерны для клеточных эукариот, к которым относятся все высшие организмы (в том числе высшие водоросли и простейшие). Возникновение эукариотической организации было одним из важнейших шагов органической эволюции. Видимо, оно открыло путь быстрой эволюции и дифференциации таксонов одноклеточных и многоклеточных. Шопф [184] описал несколько остатков эукариотических форм (возможно, это были высшие водоросли и грибы) из формации Биттер-Спрингс в Австралии (около 1 млрд. лет; фиг. 148). Отсутствие подобных форм в сходных породах формации Ганфлинт и других изученных разрезах среднего докембрия привело Шопфа к мысли, что переход от прокариотов к эукариотам мог произойти не раньше позднего докембрия.
О том, как появились высшие (эукариотические) водоросли и примитивные многоклеточные животные, палеонтологическая летопись пока дает очень мало сведений. Древнейшие известные фауны многоклеточных животных — фауна Эдиакары в Австралии (фото XXXIV) и сходные фауны с других континентов. Фауна Эдиакары состояла, по-видимому, только из мягкотелых животных, часть которых можно отнести к кишечнополостным, аннелидам и иглокожим (или сравнить с ними). Возраст фауны Эдиакары известен недостаточно точно, но породы, вмещающие ее, древнее тех, в которых найдены первые известные трилобиты, и моложе пород, возраст которых, определенный радиометрическим методом, составляет 700 млн. лет. Практическое отсутствие следов передвижения организмов, ходов или других следов жизнедеятельности в докембрийских породах заставляет предполагать, что до позднего докембрия многоклеточная жизнь не была распространена- Видимо, она возникла вскоре после перехода от прокариотов к эукариотам и быстро достигла большого разнообразия.
Предполагается, что в первичной атмосфере Земли не была кислорода, а его накопление — главным образом результат фотосинтезирующей деятельности растений. Беркнер и Маршалл [И] предположили, что содержание кислорода в атмосфере достигло уровня, при котором возможна жизнь клеточных животных (около 1% содержания кислорода в современной атмосфере), только в начале кембрия. Фишер [61] предположил, что в конце докембрия в водной среде, в тех участках, на которых развивалось много растений, создавались местные «кислородные оазисы», в которых могла происходить эволюция многоклеточных морских животных, когда содержание кислорода в атмосфере на поверхности Земли было еще недостаточным для развития здесь клеточной животной жизни.
![Фиг. 148. Glenobotrydion aenigmatis Schopf из формации Биттер-Спрингс (докембрий, Австралия) [184]. Возможно, видны стадии клеточного деления. Как полагают, темные тельца (ч) могут быть остатками клеточных ядер](/images/osnpal/1-153.jpg "Фиг. 148. Glenobotrydion aenigmatis Schopf из формации Биттер-Спрингс (докембрий, Австралия) [184]. Возможно, видны стадии клеточного деления. Как полагают, темные тельца (ч) могут быть остатками клеточных ядер")
Фиг. 148. Glenobotrydion aenigmatis Schopf из формации Биттер-Спрингс (докембрий, Австралия) [184]. Возможно, видны стадии клеточного деления. Как полагают, темные тельца (ч) могут быть остатками клеточных ядер
<< [1] ... [58] [59] [60] [61] [62] [63] [64] [65] [66] [67] [68] [69] ... [76] >>