
Разделы
- Главная страница
- Новости
- Краткий исторический экскурс
- Эра динозавров
- Гигантские растительноядные динозавры
- Устрашающие хищные динозавры
- Удивительные птиценогие динозавры
- Вооруженные рогами, шипами и панцирями
- Характерные признаки динозавров
- Загадка гибели динозавров
- Публикации
- Интересные ссылки
- Статьи
- Архив
Статистика
Д. Рауп, С. Стэнли. «Основы Палеонтологии»
![Фиг. 122. Процентное содержание современных видов моллюсков в олигоценовых и неогеновых отложениях Венесуэлы [123, 226, 227]](/images/osnpal/1-122.jpg "Фиг. 122. Процентное содержание современных видов моллюсков в олигоценовых и неогеновых отложениях Венесуэлы [123, 226, 227]")
Фиг. 122. Процентное содержание современных видов моллюсков в олигоценовых и неогеновых отложениях Венесуэлы [123, 226, 227]
Как сказано в гл. I, Симпсон [196] рассчитал, что средняя продолжительность существования вида животных составляет примерно 2,75 млн, лет. Эта величина, по мнению Симпсона, может колебаться от 0,5 до 5 млн. лет. Маркс [123] и Вейсборд [226, 227] показали, что многие современные виды моллюсков из Венесуэлы существуют 10 млн. лет и более, а некоторые виды — даже по 20—30 млн. лет (фиг. 122). Продолжительность существования многих ископаемых видов, как полагают, значительно превышает симпсоновскую среднюю оценку.
ЧАСТОТА ПОЯВЛЕНИЯ НОВЫХ ТАКСОНОВ
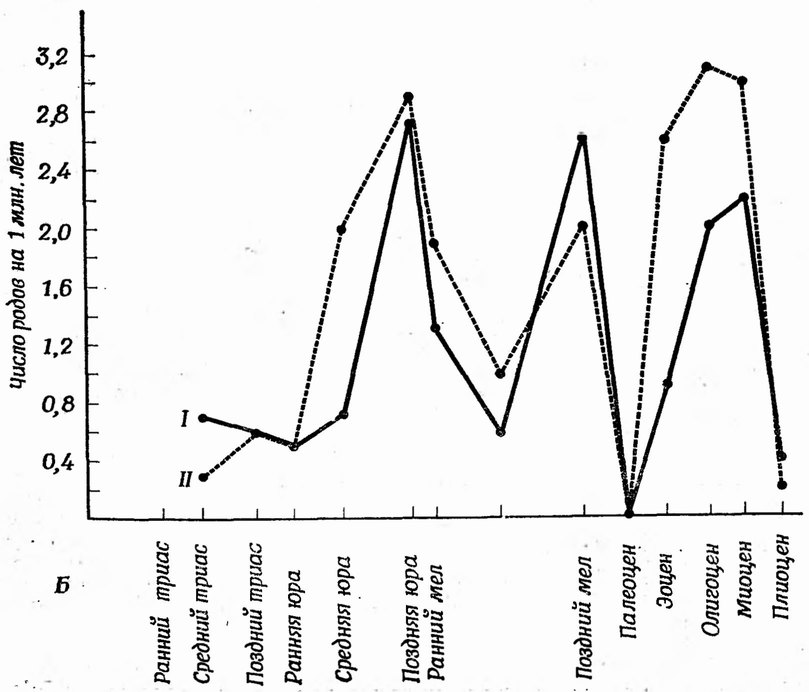
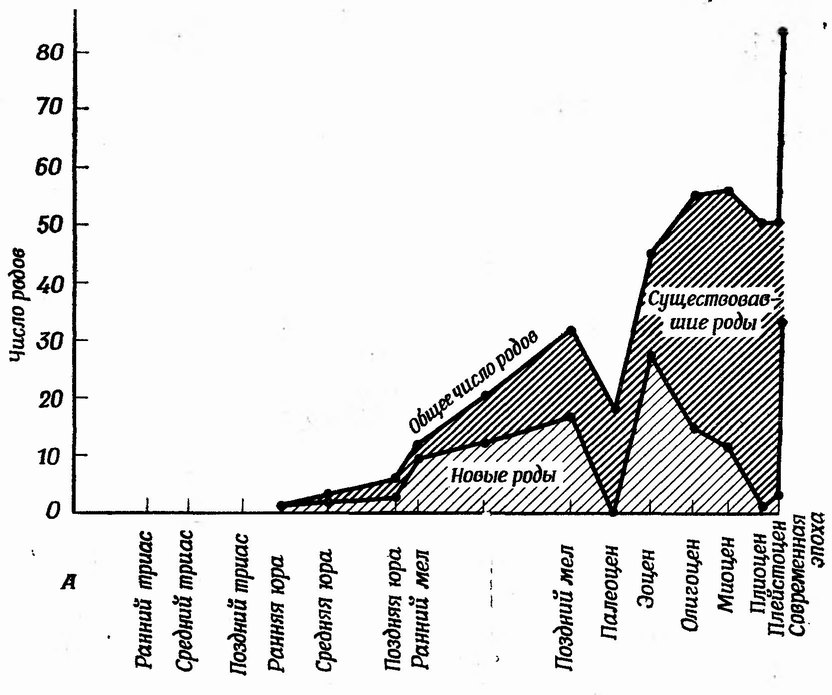
Скорость эволюции очень часто измеряется по частоте появления новых таксонов. Мы уже приводили график числа новых родов и семейств за 1 млн. лет для склерактиний (шестилучевые кораллы) (фиг. 114). На фиг. 123, А показан график числа родов склерактиний в разные эпохи мезозоя и кайнозоя; на нем выделены новые роды. По этому графику можно рассчитать скорость эволюции. Та же информация представлена в другом виде на фиг. 123, Б, где отдельно показаны появившиеся и вымершие роды. Эти две кривые очень сходны; они имеют по три пика, означающие в каждом случае три поворота р эволюции склерактиний. Возможны графики других типов. Выбор типа графика зависит от того, на что надо обратить особое внимание — на скорость увеличения разнообразия или на скорость вымирания.
Склер актинии делятся на две экологические группы — герматипные и агерматипные формы (стр. 215); поэтому графики, приведенные на фиг. 114 и 123, представляют собой упрощение. Многие семейства состоят только из герматипных или только агерматипных родов, но есть и смешанные семейства. По-видимому, эволюция герматипных родов должна отражать изменения климата или других условий среды, влиявших на рост рифов, тогда как эволюция агерматипных кораллов, менее ограниченных экологически, должна быть в целом менее зависима от изменений среды.
На фиг. 124 приведены графики, характеризующие скорости эволюции родов герматипных и агерматипных кораллов. Герматипные кораллы появились раньше агерматипных, и число герматипных родов сильно увеличилось задолго до того, как стало повышаться разнообразие агерматипных групп. Действительно, пик в позднемеловую эпоху на фиг. 123 обусловлен почти исключительно развитием герматипных родов.
Если отвлечься от кажущегося замедления эволюции в палеоцене, то число агерматипных родов с момента их появления и до наших дней постоянно возрастало (фиг. 124, А). График для герматипных кораллов далеко не столь монотонен, здесь имеются три пика разнообразия, совпадающие с пиками на фиг. 123. Стратиграфическая летопись свидетельствует о том, что первые два пика совпадают с периодами интенсивного рифообразования, от которых осталось много окаменелостей и которые за короткий срок привели к увеличению разнообразия кораллов.

Фиг. 123
![Фиг. 124. Развитие родов внутри двух крупных экологических групп Scleractinia [229]](/images/osnpal/1-125.jpg "Фиг. 124. Развитие родов внутри двух крупных экологических групп Scleractinia [229]")
Фиг. 124. Развитие родов внутри двух крупных экологических групп Scleractinia [229]
На фиг. 123 доминирующие тенденции эволюции герматипных Scleractinia сильно затемняют ход развития агерматипных родов, хотя число герматипных родов превосходит число агерматипных менее чем в два раза. Конечно, для интерпретации частот появления новых таксонов в крупных группах животных очень важен выбор включаемых в эти группы таксономических подразделений.
При интерпретации графиков следует учитывать также неполноту и искаженность палеонтологической летописи. Практическое отсутствие данных о сроках появления и вымирания родов в палеоцене, очевидно, связано со скудостью морской осадочной летописи этой очень непродолжительной эпохи. Вероятно, некоторые вымирания, отнесенные к позднему мелу, и появления новых таксонов, отнесенные к эоцену, на самом деле имели место в палеоцене. Другое любопытное явление — кажущееся увеличение числа родов кораллов после плейстоцена (фиг. 124), а дело здесь главным образом в том, что рецентные фауны известны нам лучше, чем плейстоценовые. Многие из вымерших герматипных кораллов плейстоцена, найденных в рифах, подвергавшихся действию прибоя, очень плохо сохранились и не поддаются определению. Многие агерматипные роды приурочены к глубоководным районам, и потому большая доля плейстоценовых родов может находиться в отложениях, недоступных палеонтологу. Наконец, не исключено, что немалое число родов, относящихся к обеим экологическим группам, хорошо сохранилось, и притом в доступных для исследования областях, но они пока не открыты.
Филетические тенденции
Тенденцией (trend) мы назовем направление адаптивных изменений в эволюционном ряду. При изучении тенденций проблема выделения истинных эволюционных рядов стоит не так остро, как при изучении скоростей эволюции, поскольку во многих случаях тенденции можно определить даже по довольно отрывочным палеонтологическим данным.
ОРТОГЕНЕЗ
В прошлом некоторые исследователи выдвигали концепцию ортогенеза, т. е. филетической эволюции, идущей в некоем постоянном направлении. Иногда говорят, что ортогенез-1—.это прямолинейная эволюция. Примером может служить постоянное увеличение размеров в ходе эволюции. Некоторые исследователи считали, что ортогенетическая эволюция, однажды начавшись, продолжает идти дальше в одном и том же направлении, даже если соответствующее изменение перестало быть приспособительным и может в конце концов привести к вымиранию ряда. Эта идея несовместима с концепцией приспособительных изменений под действием естественного отбора.
![Фиг. 125. Megaloceras—гигантский ископаемый олень плейстоцена; размах рогов около 2,5 м [167]](/images/osnpal/1-127.jpg "Фиг. 125. Megaloceras—гигантский ископаемый олень плейстоцена; размах рогов около 2,5 м [167]")
Фиг. 125. Megaloceras—гигантский ископаемый олень плейстоцена; размах рогов около 2,5 м [167]
Классический пример, часто цитировавшийся сторонниками теории ортогенеза, — эволюция рода Megaloceras, к которому относился гигантский олень, живший в плиоцене и плейстоцене. Рога этого животного были огромны даже для его громадного тела (фиг. 125). Рога ежегодно сбрасывались и вырастали снова, что требовало больших затрат энергии. В ходе эволюции рода размер тела возрастал, но размер рогов возрастал гораздо быстрее. Ранее предполагали, что тенденция к увеличению рогов снижала приспособленность и привела к вымиранию рода. Джулиан Гексли [86] показал, что увеличение относительного размера рогов в процессе онтогенеза характерно и для современного благородного оленя. Поскольку это увеличение в общем аллометрично (фиг. 126), Гексли предположил, что увеличение размера рогов в эволюции гигантского оленя могло происходить подобным же образом. Если размер тела и размер рогов были в эволюции генетически связаны, то давление отбора в направлении увеличения размера тела одновременно могло приводить и к увеличению размера рогов, которое могло быть либо полезным, либо вредным. Если в какой-то момент увеличение размера рогов стало вредным, то эволюция в направлении увеличения рогов могла прекратиться лишь после того, как давление отбора, направленное на увеличение размеров тела, уравновесилось давлением отбора против увеличения рогов. Нет никаких данных, которые заставили бы предположить, что под влиянием некоего таинственного «ортогенетического механизма» комплекс тело — рога перешагнул за это равновесие и дальнейшее увеличение размеров рогов стало отрицательно сказываться на приспособленности. Но Гексли подчеркивает следующее: если размер организма, например Megaloceras, увеличивается до того, что оказывается на грани допустимого, то это может привести к слишком сильной специализации и любое изменение условий среды нарушит баланс и приведет к вымиранию таксона. В быстро сменявшихся климатических зонах и биотических районах плейстоцена род Megaloceras мог погибнуть именно таким образом.
![Фиг. 126. Логарифмический график, показывающий аллометрическую связь между весом рогов и весом тела благородного оленя (Cervus elaphus) [86]](/images/osnpal/1-128.jpg "Фиг. 126. Логарифмический график, показывающий аллометрическую связь между весом рогов и весом тела благородного оленя (Cervus elaphus) [86]")
Фиг. 126. Логарифмический график, показывающий аллометрическую связь между весом рогов и весом тела благородного оленя (Cervus elaphus) [86]
Предполагают также, что аллометрический рост рогов у Меgaloceras приводил к их увеличению до гигантских размеров только у старых животных; таким образом, размеры рогов моглИ оказаться вредными у старых самцов, которые уже не участвовали в размножении из-за физиологического старения или поражений в боях с молодыми самцами. Тогда отрицательное приспособительное значение рогов могло никак не сказываться на эволюции.
Некоторые исследователи предполагали, что завернутость раковины грифеидных устриц возрастала до тех пор, пока это не стало препятствием для их открывания. Это и привело к вымиранию грифеид. Огромные размеры клыков саблезубых кошек кайнозоя также выдвигались в качестве причины вымирания тих животных. Против этих гипотез можно привести те же возражения, что и против гипотезы об «антиприспособительном» характере рогов гигантского оленя.
<< [1] ... [50] [51] [52] [53] [54] [55] [56] [57] [58] [59] [60] [61] ... [76] >>